






この記事は、社会人になったのち自然を学ぶため大学に入りなおしたわたしの卒業論文である。学会誌などには掲載していないため、こちらで公開することとした。わたしに自然史、日本の自然を教えてくださった故沖津進先生にこのブログを捧げる。


はじめに
植生を大きいスケールでとらえると、水平・垂直的な森林限界は吉良(吉良1948)の温量指数(WI)15℃・月の線に一致することはよく知られている。これは森林限界を線としてとらえたものである。しかしながら実際の森林限界や樹木限界は、連続・密閉した森林から不連続な疎林を経て裸地へと移り変わる移行帯である。横山(1995)は、こうした移行帯では、気温、積雪、卓越風、地形などが複雑に関与して形成されていることを示唆している。地域によってその要因の作用構造や程度が異なるため、地域による相違は、個々の場所で植生遷移の状態をよく分析することが重要だと指摘している。日本の中部山岳以北の森林限界の特徴として、沖津(1984、1985)は、森林からハイマツ(Pinus densiflora (Pallas) Regel)帯へ急激に移行すること、温量指数的には森林の成立が可能な標高であっても、強風や多雪によって森林が排除されることを指摘している。これは日本の山岳は十分な高さを持たないため、温量指数15℃・月に相当する標高が、山頂や稜線付近の特殊な気象条件が支配する範囲に入っており、森林の成立が阻まれていると考えられる。この空白域にハイマツ帯が成立している。日本の山岳地帯では亜高山帯構成樹種が、矮生型をとって匍匐する樹木分布の上限や高木限界に対応する現象が乏しい(沖津 1984)。
これらの条件に対する例外が、十分な標高をもち、ハイマツが分布しない富士山である。富士山は噴火の年代が新しいため、植生遷移の途上にある(大沢ら1971)。温量指数15℃・月相当の標高(約2900m)までカラマツ高木が到達しているのは、西斜面だけである。森林構成種が高木になることを制限され、矮生型をとることで低木化しており、その形状は現在の環境条件や森林帯形成の歴史を反映していると考えられる。
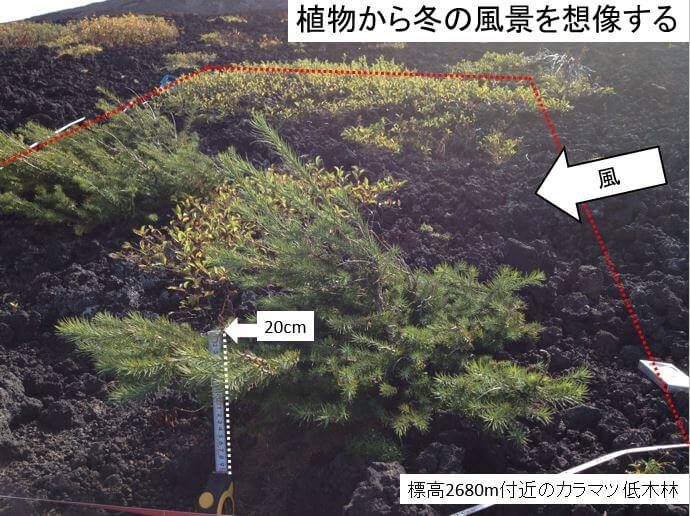
本研究では、移行帯に関する用語は岡(1992)にならい、森林構成種が高木で連続的に生育できる限界を森林限界、森林構成種そのものが矮生化し生育している生育限界を樹木限界として区分する。森林限界と樹木限界をあわせて樹木限界移行帯として扱う。 富士山の現在の樹木限界移行帯の標高や範囲は斜面によって異なるが、移行帯内部では、標高が低くなると偏形の程度が軽くなる(岡1980)。偏形は冬季に吹きつける雪氷片や細かい砂礫で樹皮が傷つき、枝の含水量が致死レベル以下に低下し枯死することで形成されている(Maruta 1996)。富士山南斜面では、最初に矮生型カラマツが定着し、その山頂側に新たな矮生型カラマツが定着することで冬季の環境が緩和され、下方に分布しているカラマツの幹が立ち上がり始め、それが繰り返され、標高が下がるに従い偏形化が軽減していた(丸田ら 2009)。富士山で最も樹木限界移行帯が温量指数15℃・月に対応している西斜面でも、同じように標高に対応した偏形タイプの変化が確認されている(岡1980; 岡1990)。本研究では、富士山北西斜面風衝地の矮生化したカラマツ林に着目し、植生構造からカラマツ低木林立地の規定要因と動態を明らかにする。
調査地概要
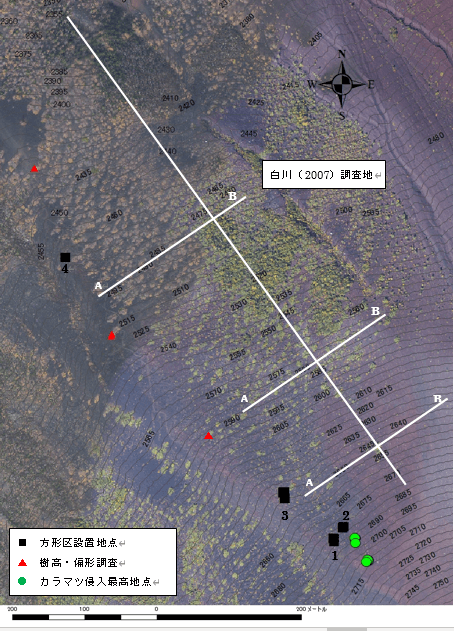
調査地域は富士山山梨県側5合目の御庭上部の、北西斜面2400m~2700mである。御庭寄生火口列が斜面の西側と東側にあり、標高2450m付近を境にして、斜面の下方は御庭第一溶岩流によって約1600年前に、上方は旧期溶岩流によって約2200~3200年前に形成された(津屋1971)。気候は、太平洋型気候を示し、冬季の積雪は少ない。富士山頂に気象観測所が設置されていたが、1999年にレーダー観測が廃止され風向風速が観測されなくなった。2004年からは無人化し、夏季のみ観測を行っている。富士山頂の年平均気温は-6.4℃(1974~2003年)、年平均気温は11.4m/s(1974~1999年)、夏季の卓越風向は西、冬季は西北西である。富士山頂の温量指数は1.2℃・月であり、温量指数15℃・月の高度は2930mである(岡1991)。本調査地である北西斜面で気象観測を行った岡(2000)によると、標高2685mで年平均気温は1.1℃、最寒月で月平均-12.7℃、土壌凍結期間は12月15日から3月16日であった。標高2800mでは年平均気温が0.3℃、最寒月で月平均-13.2℃、土壌凍結期間が11月23日から4月23日であった。 白川(2007)は、本調査地から150m西側の矮生型カラマツ林内でベルトトランセクトを設置し調査を行っている(図10)。図中の白い線が白川(2007)の調査測線である。A、Bは図11の断面図と対応している。本研究では白川(2007)が調査した場所の南西縁で尾根に沿って調査地を設置し、移行帯内でも更に風衝地に立地している矮生型カラマツがどのような動態をしているのかを検討した。
方法
1.空中写真判読と数値地図5mメッシュの図化
調査対象地域の樹木限界移行帯の変化を確認するために空中写真(米軍撮影、国土地理院撮影、山梨県撮影)の判読を行った。1948年、1975年、2008年の写真を用いた。1948年と1975年撮影の写真は、フィルムを印画紙に密着焼き付けしたものをスキャナでデジタル画像化した。それを幾何補正し、オルソ画像を作成した。使用したソフトはアジア航測株式会社ADIMS:計測名人である。補正の精度は基準点の誤差が数値地図1:25000の位置に対して5m以内である。2008年撮影の写真は、すでに幾何補正まで済んだデジタルデータを提供していただいた。得られたオルソ画像をGISソフトウェア(ESRI社 ArcGIS)に取込み、樹木限界移行帯の南西縁に位置するカラマツパッチ群落の位置とサイズを読み取った。現地でパッチの直径サイズ1mを越えるものは、読み取り可能であった。また、ミヤマヤナギ(Salix reinii Franch. et Savat.)やイタドリ(Fallopia japonica (Houtt.) Ronse Decr. var. japonica)もパッチを形成するので、カラマツであることを現地で確認してハンディGPSを用いて観測し空中写真にプロットすることで他種と区別した。
2.植生調査
白川(2007)の調査地の南西縁で尾根に沿って巾10mのベルトトランセクト(標高2426m-2710m)を設置した。ベルト内で出現するパッチ群落について偏形タイプ、樹高、球果の有無、稚樹の有無を記録した。本調査地で優占するカラマツはその偏形化の程度によって丸田(2009)にならい以下の4種に分類した。
①矮生型;幹は鉛直に伸長できず、幹や枝は地表を這ってマット状の樹形となる。
②立上り型;矮生型と同様にマット状に広がるが、その中から数本の幹が風下側に斜めに伸長する。立上った枝の風上側には枝がない。矮生型より標高の低いところや風下側に分布している。
③旗型;立上り型よりも、さらに下方に分布し立上った幹がすぐに鉛直に伸長、もしくはマットから立上ったすぐには風下側に斜上するがその後鉛直に伸長する。風上側に枝がなく、旗に似た形になる。マット状の部分が残り、地表付近が枯れて白骨化していることが多い。
④先端対称型;旗型とほぼ同じ樹型であるが、幹の先端部分では、風上側にも枝が伸び、正常な幹となる。マット状の部分は、さらに枯れが進み、白骨化した部分が多くなる。
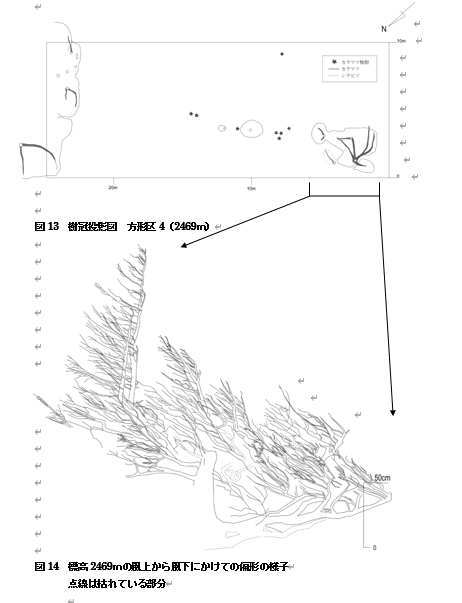
樹高は、矮生型カラマツでは、カラマツ数個体が複雑にからみあい、ひとつのパッチを形成しているものが多く、その場合はパッチ群落表面の最も高い部分と最も低い部分を樹高として記録した。樹高50cm以下で、当年生を除くカラマツ個体を稚樹として扱った。トランセクト内に、偏形タイプが多様でパッチサイズが大きかった標高2469mで10×25mの方形区を設置した(方形区4)。また矮生型が優占し1948年の空中写真でパッチの存在が確認できたもののなかで、高標高域に位置する場所に7×8mの方形区を設置した(方形区3)
結果
樹木限界移行帯の変化
調査地の詳細な地形図を図2に、カラマツ落葉前の様子を図3、積雪期の様子を図4に示す。
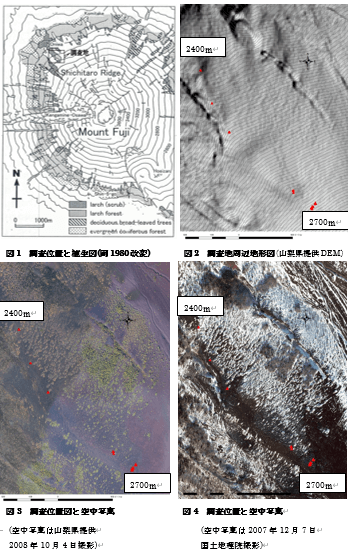
尾根部は植生が欠如し裸地になっている様子がこれらを比較するとよくわかる。裸地になっている部分(2500-2600m付近)でふたつの小山があることも確認できる。裸地の尾根から北東にかけてパッチ状に植生が散在し、積雪期にはそこに雪がたまり独特の模様をつくりだしている。数十年単位ではあるが、時代変化をみるために、1948年、1975年、2008年の空中写真を用いて調査地の尾根部のパッチ長径の変化を測定した(図5)。標高2660m以下では1948年からパッチ群落が確認できた。変化は小さいながらも長径が増えているパッチも存在した。2668m以上の高標高域では2008年でのみパッチが確認できた。
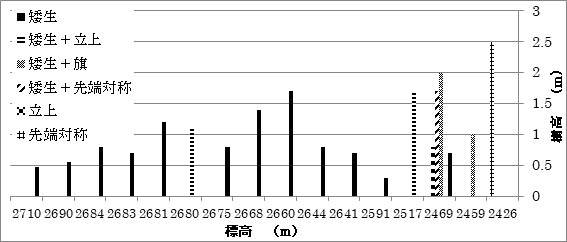
各樹型と樹高の分布
カラマツの樹型タイプの変化を標高ごとに模式図であらわした(図6)。

高標高域では矮生型のカラマツが分布し、風衝地形が著しい場所で裸地が残り、その後標高が下がるに従い、立上り型、旗型、先端対称型と偏形が軽減していた。標高と樹高の関係を示したのが図7である。パッチ群落の樹高差がある場合、最も高い部分をこの図では樹高としている。2675m以上の標高では、樹高が1m前後となっており、偏形タイプは矮生型が優占していた。標高2469m以下では、樹高が高くなり偏形タイプも立上り型や旗型が出現していた。森林限界付近である2426mでは樹高は2mを超え、偏形タイプも先端対称型になっていた。
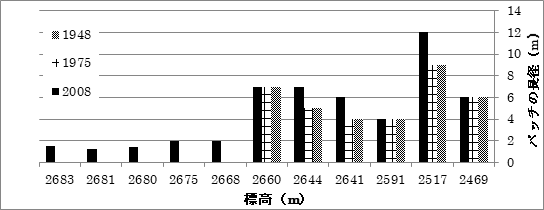
ベルトトランセクトの稚樹と球果
ベルトトランセクトの球果の有無を表1に示す。2684mでカラマツの稚樹が2個体確認された。尾根部周辺では2684mが稚樹定着をしている最高標高だった。カラマツは球果をつけて種子を散布した後、数年間は球果を枝上にそのままつけているので、種子生産の有無を判定することができる。稚樹定着が確認できた2684mでは、球果の生産が行われていた。樹木限界の2710mと2690mの2個体はまったく球果をつけていなかった。そこから標高2660mまで種子生産は確認されなかった。
方形区
設置した方形区で樹冠投影図を描いた(図12、13)。
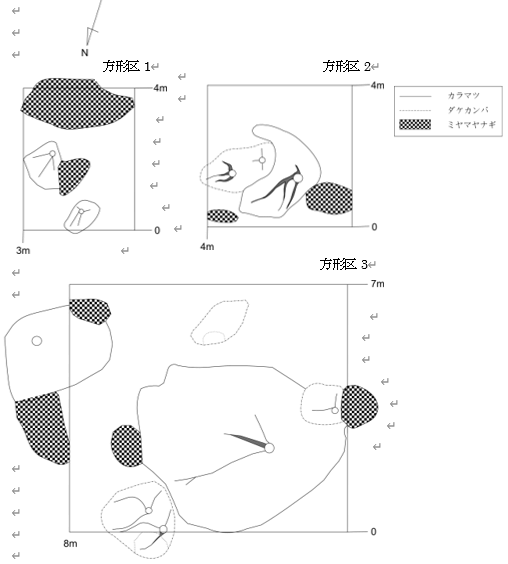
方形区1(2683m)、方形区2(2680m)、方形区3(2644m)
本調査地でパッチ群落を形成している樹木限界に近い位置に設置した方形区が1と2である。また、パッチ群落としては大型のもので1948年の空中写真でパッチの存在が確認できたものが方形区3である(図12)。実線がカラマツで、破線がダケカンバ(Betula ermanii Cham.)、網掛けがミヤマヤナギを表す。丸い白抜きが確認できた主幹で、そこから延びる線が枝の伸長方向である。方形区2と方形区3ではダケカンバが分布していた。また方形区1-3で、カラマツとミヤマヤナギが共存していた。2683mに設置した方形区1では、ミヤマヤナギの分布面積のほうがカラマツより大きかった。方形区4は偏形が軽減されタイプが多様になっていた箇所に設置した(図13)。実線がカラマツ、破線がシラビソ(Abies mariesii Masters)を表す。また、パッチ群落とパッチ群落の間の裸地に稚樹が定着していた。各方形区の毎木調査結果を表2に示す。方形区1では、カラマツはパッチ内に2個体あり、どちらも樹高は1m未満であった。方形区2では、カラマツが1個体、カラマツより樹高が低いダケカンバが2個体生育していた。方形区3では、主幹の地際直径が10cmの大きな矮生型カラマツが生育していた。カラマツより樹高の低いダケカンバが、大きなカラマツ個体周囲に4個体分布していた。方形区4では、カラマツの地際直径が大きく樹高が高くになるにつれ偏形タイプが旗型や先端対称型になっていることがわかる。定着していた稚樹は、5cmから50cmまでほぼすべての樹高を持つ稚樹が存在していた。方形区内にシラビソが1個体分布していたが、シラビソの稚樹は確認できなかった。
伸長生長できないカラマツ
標高2600mを越えると樹高が約1m前後だった(図7)。
方形区3を設置した標高2640mでは、ある一定の高さ以上に伸びた枝が白く枯れているのが確認できた(図9)。

標高2469mでは、南西から北東にかけて偏形タイプが立上りから旗型へと変化していた。地表周辺では枯損して白骨化している部分も確認できた(図14)。

点線は枯れている部分
斜面下から山頂側を見上げるようにして樹木限界の2690mと2710mでカラマツを撮影した写真がそれぞれ図16と図17である。写真の右側が南西の尾根で左側が北東である。両個体共、樹高約50cmだったが、すでに偏形し地表面を這うような形をしていた。


考察
本研究で設定した調査地は森林限界移行帯上部に位置し、山頂が近くなるに従い生育環境が厳しくなるため、樹木の定着は困難で、最も偏形化した樹形である矮生型になる。そして、標高が下がるにつれて気候が緩和され、偏形化の程度が軽くなる。本調査地の北西斜面でも同様に(図5)。標高が下がると偏形が軽減し、偏形の軽減に伴い樹高が高くなっていた。(図6)
加えて本調査地では標高の高低差だけでなく、南西から北東の方向にも偏形度の違いが確認されている(白川2007)。白川(2007)は、本調査地から150m西側の矮生型カラマツ林内での調査から、尾根部の南西から谷地形の北東に向かってカラマツの偏形度の変化を実証している。富士山では11月~4月にかけて西~北西の強風が続き、独立峰である富士山は、その風が山体に沿って流れることが知られている。太平洋型気候により冬季の積雪量は少なく、特に風衝地である本調査地(岡 1980)は1月頃まで砂礫が露出している。そのため、雪氷片だけでなく細かい砂礫も矮生木限界のカラマツに吹きつけ、枝を傷つける。矮生木では、毎年初夏に新しいシュートが伸びても、冬にはこのようにして枯死することを繰り返しており、幹が鉛直に立ち上がることができない。積雪で保護される部分より上部に突き出ていたと考えられる枝が、枯損していた。本調査地は、冬季に最も風上になる尾根に位置している。カラマツ矮生木は風に吹き飛ばされた砂礫をほとんど捉え、それに保護される位置にあるカラマツを、損傷から守る(Maruta 1996)。そのような環境の緩和作用を受け、尾根から谷にかけて徐々に樹高が立ち上がっている。それが繰り返されることにより、南西から北東にかけて地形とカラマツパッチの環境緩和作用がより強く働き、偏形が軽減され樹高が高くなっていくと考えられる。
以上のように、標高による上下の植生の対応がみられるだけでなく、冬季卓越風の方向に対応した植生の反応が観察された。そして、調査地ではカラマツ矮生木がパッチ状に散在している。このパッチ内でもカラマツの防風作用がはたらいていた。その方向は山頂側から下に向かうのではなく、冬季卓越風の風上から風下に向かっていた。富士山のカラマツ偏形の変化は冬季の環境に対応しており(丸田2009)、パッチ内でも環境の厳しい風上の先端に位置するカラマツは、冬季の強い季節風を受けて幹の鉛直方向の生長ができないが、自身で風を弱める働きをすることで、パッチ内で風下側に位置するカラマツの伸長生長を助けていることが考えられる。調査地内で比較的低い標高域である2469mでも、風上側にあるカラマツは伸長成長ができていなかった。2469mで設置した方形区4では、5cmから50cmまでほぼすべての樹高を持つ稚樹が存在しており、ここはカラマツの稚樹の定着・成長にとって適した環境であることが示唆される。カラマツの稚樹定着位置は、偏形が最も大きいカラマツの風下側に位置していたことから、今後これらが生長していきパッチの構成員になり、パッチを拡大することで風下側のカラマツの伸長生長を促していくことが予測できる。その拡大は冬季卓越風の風下側を埋めるように進んでいくと考えられる。パッチの拡大方向について高橋ら(1994)は大雪山の高山風衝砂礫地で、パッチ状に分布する植生の島が、積雪分布と相関関係にあることを実証した。また、丸田(未発表)は富士山南斜面の2450mから2500mで、樹高1m以下の矮生型カラマツが冬の間積雪に覆われることを確認している。先に述べたように本調査地もパッチ状にカラマツ矮生木が散在しており、冬季卓越風により同一方向に島が拡大している。これらのパッチ群落は冬の間、積雪に覆われて保護されることが考えられる。2600m前後にあるパッチやカラマツ侵入最高標高地点(2710m)では、カラマツがミヤマヤナギと連続したパッチを形成し樹高は1m以下である。カラマツ自身による風の防護作用が働かない樹木限界移行帯上端ではミヤマヤナギとパッチを形成できた個体が、厳しい冬を生き延びていることがわかる。パッチの発達方向と積雪域がきわめて整合性が高いことから、冬季卓越風に対しミヤマヤナギやカラマツ自身により北東方向へしっぽ状の積雪域が形成され、その保護下にパッチが発達していることが考えられる。
本調査地の矮生型カラマツの分布上限は、2710mであった。個体数は少なく、サイズも小さかった。カラマツはまだ種子生産をしておらず、周囲に定着している実生もみられなかった。しかし、すでに風衝形をとって強風に対応しており、今後も矮生型カラマツへと生長していくことが可能であると考えられる。このように矮生型カラマツが環境を緩和し、パッチ面積を増大させることにより植被率増加に寄与し、風下のカラマツの伸長成長や後に続く亜高山帯針葉樹種の定着を可能にさせていると考えられる。本研究の対象地である北西斜面尾根部の樹木限界移行帯上部では、山体に沿って吹き込む冬季卓越風の風向と積雪が重要な環境要因であることがわかった。2469mはカラマツの稚樹の定着・成長にとって適した環境であることが示唆される。
カラマツとダケカンバとミヤマヤナギは、先駆樹種といわれているが、ここの森林限界移行帯上部のように強風が樹木定着の阻害要因となっているところでは、まずミヤマヤナギが定着し、カラマツが定着、その後にダケカンバが侵入するということが示された。このような順序は富士山では一般的である。シラビソは2469mでのみ1本確認された。共存するカラマツよりも樹高は若干低いが、ほぼ同じ高さにまで達している。このことから、すでにカラマツ優占する先駆林から、遷移の次のステージに移行を始めている段階といえる。しかし、まだシラビソが生息している風下側の裸地にはカラマツ稚樹が定着していることから、安定しているパッチのそばから部分的に遷移が移行していく様子が示された。
コメント