







はじめに
日本列島は大陸の東縁部に南北3000㎞にわたり、大小あわせて6800を越す島々で形成されている(出典:公益財団法人日本海事広報協会)。気候は亜熱帯の沖縄から亜寒帯の北海道まで網羅する。標高は最も高い地点が3776mである。また、日本海側では冬に雪が多く、太平洋側では夏に雨が多い。それに応じ自然環境も多様である。
植生(Vegetation)とは森林や草原など緑の広がりを示す概念である。同じような立地には同じような植物群落(Plant Community)がみられるため、立地条件や植物種の組成で類型化されている。植物群落は植生の単位として使われる。植生図は植物群落とその類型を地図として表したものである。
似た用語に植物相(Flora)がある。植物相は地域に生育する全ての植物を同定して、種名などを記した種のリストで表す。
本記事ではにっぽんの森林を植生や植物相により概観することを目的とする。
植生変遷史
にっぽんの植生と植物相は、第三紀から第四紀にかけて形成された。第四紀へ移行したのは約260万年前である。日本列島は温暖で気候変動の少ない第三紀から、氷期が周期的におとずれた第四紀へ移行した。第三紀に繁茂していた植物相は特定の時期、特に寒冷化が進んだ時期(約300~250万年前、約120~90万年前、約40万年前以降)に集中して絶滅した。第四紀に繁茂する植物相は約140~50万年前にかけて徐々に出現した。
80万年前以降、約10万年周期で氷期と間氷期が繰り返され、少なくとも4回(ギュンツ、ミンデル、リス、ウルム)の大きな氷期と間氷期の繰り返しがあった。氷期と間氷期の繰り返しの中で、南方系、北方系の植物の渡来、日本列島内での南北、上下方向への移動、列島内での独自の進化と絶滅があり、日本の植物相と植生が形成されてきた。例えば、ミンデル氷期にはグイマツ、チョウセンゴヨウ、シラビソ、トウヒなどの亜寒帯性の針葉樹が数多く見られるのに対して、ミンデル-リス間氷期にはアデク、ヤマモモ、ナギ、ツクバネガシなど温暖型の常緑樹が出現する。
清水善和2014「日本列島における森林の成立過程と植生帯のとらえ方ー東アジアの視点から」地域学研究第27号 P27
日本全国の花粉分析から描き出された日本列島(関東中心)の最近12万年におよぶ植生の変遷を述べる。まず,最終間氷期(リス–ウルム間氷期:約12万年前)は夏冷涼・冬温暖な気候で,シマサルスベリ,センダン,ナンキンハゼ,アブラギリ,アカガシ亜属のような暖温帯性の広葉樹が多かったが,最終氷期(ウルム氷期)の前半(12~6万年前)になると冷涼・湿潤な気候に変わり,日本中でスギ林が目立つようになった。中頃(6~2万年前)には寒冷・乾燥の気候となり,スギは衰退してナラ類やハンノキ,カバノキなどの冷温帯性落葉広葉樹とトウヒ,カラマツなどの針葉樹の混交する森林が成立した。最終氷期最盛期(2–1.5万年前)には,現在より平均して気温が5~7°Cほど下がり,植生帯は800–1200 m降下したという。
日本でも、鮮新世から更新世への植生や植物相の変遷が植物化石資料によって追跡されている。三木茂(1948)は近畿地方を中心に鮮新世以降の植物化石包含層を、1⃣メタセコイア層、2⃣ハマナツメ層、3⃣スギ層、4⃣カラマツ層、5⃣ナンキンハゼ層、6⃣ムクノキ層(沖積層)に区分した。その後、市原(1960)は大阪層群の層序を確立するにあたって、三木の化石層を層序学的に再検討し、日本からの絶滅種が消滅する層準を明らかにし、イチョウ、セコイア、フウなど日本からの絶滅種を多く含む後期鮮新世の地層をメタセコイア植物群繁栄期の地層、メタセコイアやオオバラモミが消滅する前期更新世末までをメタセコイア植物群消滅期の地層として区分した。
百原新2005「6.2 植物群と植生」-第四紀学 p191-219朝倉書店
メタセコイアのように、第三紀前半に北半球の広い地域に分布したが、第三紀の後半に分布域を縮小し、現在中国中南部に残存した裸子植物には、イチョウ、イヌカラマツ、スイショウがある。このほか、スギは日本と中国東部に残存し、コウヤマキは日本だけに残った例である。日本や中国西南部に多くの植物が絶滅をまぬがれて残存した原因として、これらの地域では氷期に大陸氷床や山岳氷河の影響を受けなかったこと、中国中部以南では山脈がほぼ南北に走っていて第四紀の激しい気候変化に応じて植物が移動しやすかったことが挙げられる。
第三紀そして第四紀を生き残った植物の子孫がいまでも存在していることを知ると、日本列島の植生に面白さを感じないだろうか。絶滅を免れた植物たちは現代にっぽんのどんな地域を好んで生きているのか?そんな視点で植生を知ると面白い。植生史はとても奥深いので、また改めて記事を書きたいと思う。
現代の植生について
にっぽんには約5800種の植物が分布し、そのおよそ30%の約1800種が日本固有種である。
百原新2005「6.2 植物群と植生」-第四紀学 p191-219朝倉書店
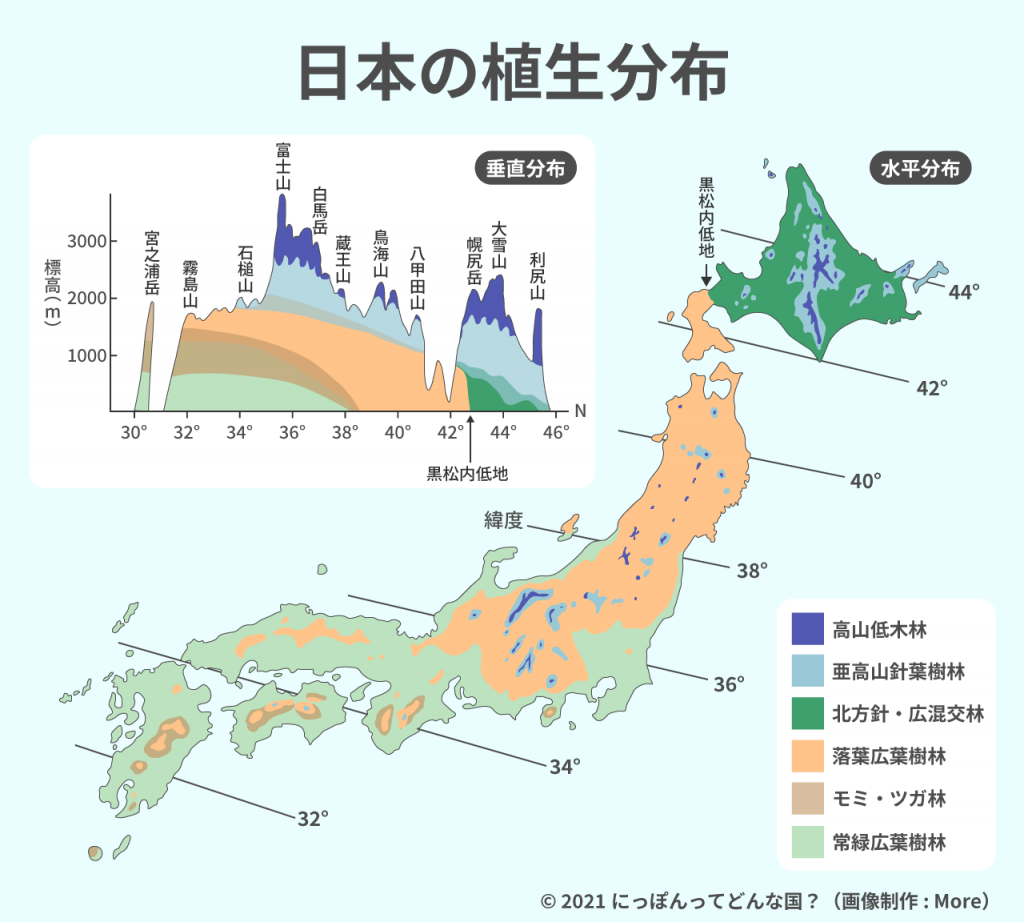
標高の違いによる植生や植物相の変化は「垂直分布」、緯度の違いによる同様の現象は「水平分布」とよばれている。どちらも温度環境の違いが主な原因と考えられている。地形変化の少ない地域では垂直分布と水平分布がほぼ対応し、緯度1度の北進と標高100mの上昇が等しいといわれている。しかし地形変化や気候変化が複雑なにっぽんではその変化が一様ではない。
日本の植生帯区分は下記の表のように研究者によって議論されてきている。植生帯区分で相違点が際立つのは2点ある。1点目は暖温帯と冷温帯の間に「中間温帯」を認め独立した植生帯とするかどうかである。2点目は垂直分布のなかで「ハイマツ帯」を高山帯に含めるか、亜高山帯上部に独立した植生帯として位置づけるかどうかである。
| 提案者 /気候帯 | 寒帯 (高山帯) | 亜寒帯 (亜高山帯) | 冷温帯 | (中間温帯) | 暖温帯 | 亜熱帯 |
| 福嶋(2017) | 常緑針葉樹林 | 落葉広葉樹林 (汎針広混交林) | 常緑針葉樹林 暖温帯落葉広葉樹林 | 常緑広葉樹林 | 常緑広葉樹林 | |
| 山中(1979) | 亜寒帯林 | 冷温帯林 | 中間温帯林 | 暖温帯林 | 亜熱帯林 | |
| 大場 (1975) | 低小草原帯 | 針葉林帯 | 温帯夏緑広葉林帯 | 亜熱帯常緑広葉林帯 (照葉樹林) | ||
| 吉岡 (1973) | 高山植生 | 亜高山(亜寒帯) 針葉樹林 | 落葉広葉樹林 (北方針・広混交林) | モミ・ツガ林 | 常緑広葉樹林 | 亜熱帯多雨林 |
| 堀川 (1968) | 高山荒原 | 山岳針葉樹林 | 落葉広葉樹林 | 中間針葉樹林 | 常緑広葉樹林 | 常緑広葉樹林 |
| 宮脇ら (1967) | 高山帯 | 亜高山針葉樹林帯 (トウヒーコケモモ クラス域) (ハイマツ帯含む) | 夏緑広葉樹林帯 (ブナクラス域) | 常緑広葉樹林帯 (ヤブツバキクラス域) | (亜熱帯) | |
| 鈴木 (1961) | 高山帯 | 亜寒帯林 | 冷温帯林 | 中間温帯林 | 暖温帯林 | 亜熱帯林 |
| 吉良 (1945) | 針葉樹林帯 | 温帯落葉樹林帯 | 暖帯落葉樹林帯 | 暖帯照葉樹林帯 | 亜熱帯多雨林 | |
| 中野 (1942) | 亜寒帯林 | 冷温帯林 | 暖温帯林 | 亜熱帯林 | ||
| 本多 (1912) | 寒帯林 | 温帯林 (ブナ帯) | 暖帯林 (亜熱帯林) (カシ帯) | 熱帯林 | ||
| 田中 (1887) | ハイマツ帯 | シラベ帯 | ブナ帯 | 間帯 | クロマツ帯 | アコウ帯 |
植生帯区分の研究者による解釈を比較する際に必要な論点をさらに細かく見ていこう。
暖かさの指数(温量指数)
植生帯区分を論じる際に重要な指標がある。それが暖かさの指数(温量指数)である。暖かさの指数は大阪市立大学名誉教授である吉良竜夫(1919-2011)により考案された。暖かさの指数WIとは、植物にとって1年間利用できる温度の積算量がどれくらいあるのか?を数値化したものである。12か月分の月平均気温データのうち、5℃以上の部分を足し合わせたものである。
| 気候帯 | 温量指数(WI) |
|---|---|
| 熱帯 | >240 |
| 亜熱帯 | 240-180 |
| 暖温帯 | 180-85 |
| 冷温帯 | 85-45 |
| 亜寒帯 | 45-15 |
| 寒帯 | <15 |
例えば、ブナの分布上限・下限は暖かさの指数が85-45ともっとも密接な関係がある。ただし、温量指数は降水量が十分にある地域にしか適用できない。
冬季の寒さも植物にとって重要な指標であり、吉良先生は寒さの指数も考案された。寒さの指数とは、月平均気温が5℃以下となる月の月平均気温から5℃を引いた値を足し合わせた数値のことである。冬に休眠しない植物は寒さ指数と対応関係がある。
中間温帯
日本の植生が論じられた初期から、暖温帯の照葉樹林帯と冷温帯の夏緑樹林帯との間に特有の植物群落が出現することが認識され、鈴木(1961)により中間温帯と命名された。
中間温帯林の定義が研究者や地域によって異なることが興味深い。研究室のゼミ旅行で中間温帯林を観察したことがある。そのときの事前資料を記事にしたのが下の「伊豆半島におけるモミ林の分布帯」である。いわゆる中間温帯林はモミ(Abies firma, 分布: 本州秋田県以西,四国,九州屋久島まで)やツガ(Tsuga sieboldii,分布: 本州福島県八溝山以西,四国,九州屋久島まで,韓国鬱陵島)が目立つ樹林が現れ、場所によってはモミ・ツガ林と呼べるような景観となる。また、地域によってはモミのほかにイヌブナやシデ類などの落葉広葉樹が混じる独特の樹林帯も存在する。

実際に見に行ったときの工程

日本の中間温帯の成立過程を考える上で、東アジア大陸部での植生配列が参考になる。
清水善和2014「日本列島における森林の成立過程と植生帯のとらえ方ー東アジアの視点から」地域学研究第27号pp19-75
-略-
照葉樹林帯(低地)と針葉樹林帯(亜高山帯)の間にどんな植生が現れるかというと,針葉樹のツガ属の優占する林(場所によってはツガ純林)やマテバシイ属(Lithocarpus)が多い常緑樹林が出現することが多い。石塚(1977)もこのような温帯性樹林の出現は,台湾,中国南部をへてヒマラヤ山脈につらなる一連の地域に共通の特性であると述べている。
ちなみに,北米東岸アパラチア山脈南部のグレート・スモーキー山でも,山地の中腹辺りで落葉樹のアメリカブナ(Fagus grandifolia),ユリノキ,アメリカフウなどと針葉樹のツガ(Tsuga canadensis)が混生しており,中間温帯的相観を呈しているのが興味深い.-略-こうした他種と混交するブナ属の在り方は,日本の中間温帯に特徴的に現れるイヌブナ(Fagus japonica)と似ている。イヌブナは岩手県北部を北限として本州,四国,九州の中間温帯域に見られるが,中部以北の太平洋側に多く,本州中・北部の日本海側には存在しない。日本海側のブナ林のような純林を作らず,モミやナラ類,シデ類を交えた混交林を構成する(野嵜 2005)。
清水善和2014「日本列島における森林の成立過程と植生帯のとらえ方ー東アジアの視点から」地域学研究第27号pp19-75
日本列島の植生を考える時に、周辺や世界がどうなっているのかを知ることも重要である。今後はにっぽんを知るために世界を引用する記事も書いていきたい。
ハイマツ帯
ハイマツ帯は日本の植生帯区分の議論で2つ目の論点である。日本では伝統的に森林限界を超えると高山帯と呼び、ハイマツ帯を高山帯に含める研究者が多い。しかし、高山帯に含めることは少し厳しいのではないか、という説とその根拠を紹介する。
日本の山は夏の気温だけから考えると、中部山岳地域で2870mくらいまで亜高山針葉樹林、3200mくらいまでハイマツに覆われる計算になる。しかし実際の森林限界は2500mくらいまで低下し、その上のハイマツ群落も分布はパッチ状で、パッチの間を埋めるように高山植物の群落ができている。このような分布が生じるのは、日本の山が冬場、世界一の強風と世界有数の多雪といった厳しい条件下にあるからである。
小泉武栄(2017)「日本の高山地域の植生」図説日本の植生第2版 朝倉書店
ハイマツ群落の生産力は森林のものとほぼ等しく、高山植物群落や寒帯ツンドラの10倍以上の値を示す。ハイマツ群落は生産力の面からは森林群落に属している。ハイマツ群落の分布域は、夏の温度条件以外の環境条件によって高木の針葉樹林がなくなる場所といえる。日本の山岳上部は冬期の偏西風が強く、世界一の強風帯である。雪の量もきわめて多い。高木だと、強風のために、冬のあいだに風上側の枝や葉が傷を受けたり、傷口やクチクラ層から強制的に水分が蒸発してしまい、その結果、乾燥で枯れてしまう。一方、ハイマツ群落には葉がぎっしりと着いているので、初冬に雪をすばやくとらえて、雪の中に埋もれる。ハイマツは低木のために幹が柔軟で、雪の圧力で折れることはない。ハイマツ群落を覆う雪は、冬のあいだ、強風による葉や枝の損傷や乾燥から群落を保護するのである。ー略ー
沖津進(2017)「ハイマツ帯の構成とハイマツ群落の分布」 図説日本の植生第2版 朝倉書店
ハイマツの分布の中心は東シベリアにあり、日本列島は南端に位置する。
東シベリアでは、ハイマツはグイマツ疎林の林床要素として分布し、単独では植生帯をつくらない。ハイマツ群落は、東シベリアに広がるグイマツ・ハイマツ林から高木性のグイマツが冬期の強風や多雪などで欠如し、林床のハイマツだけが残存したものである。したがって、ハイマツ帯は森林限界移行帯や高山帯には相当せず、下方の森林帯とは独立した別個な植生帯ととらえられる。
亜高山帯針葉樹林の上限を越えると,厳しい環境(低温,強風,岩石露出など)のために森林(高木林)を維持することができず,植生は低木林や草原に移り変わる。この森林の上限を森林限界(forest limit)とよび,森林限界より上方を一般に高山帯という。森林限界は水平分布,垂直分布の両面においてWI=15のラインに一致することが知られている(丸田 2000)森林限界から高山帯への移行の仕方はいくつかのパターンに分けることが可能だが
清水善和2014「日本列島における森林の成立過程と植生帯のとらえ方ー東アジアの視点から」地域学研究第27号pp19-75
(Tranquillini 1979; Holtmeier 2003),大きくは,亜高山帯の森林を構成する針葉樹が密度や樹高を低下させながら上昇し樹木限界(tree limit)に達する場合と,亜高山帯を構成する樹種とは異なる種が独自の低木林(または矮低木林)を形成する場合に分けられる。前者のような亜高山帯から高山帯へ移行する部分を森林限界移行帯とよび,北米のロッキー山脈やヨーロッパ・アルプスでよく見られる。日本の垂直分布では後者のタイプが主で,ハイマツがかなりの高度幅をもって帯状に出現するのが特徴である。
大陸では日本のようなハイマツ帯を形成することはほぼなく、高山の森林限界の上方でハイマツ帯と呼べるような純林状の植生帯を形成するのは日本独自の現象である。
ところで、富士山(3776m)にハイマツは存在していない。ハイマツの種子は主にホシガラスによって運ばれる。ホシガラスはハイマツの球果をもぎとると近くの見晴らしのよい場所で食べる習性があり、長距離運ばれる可能性が低いと考えられている。現在、富士山の森林限界付近では落葉性針葉樹のカラマツが低木状となり森林限界移行帯を形成している。富士山の西斜面では温量指数WI=15に近い2800mに達しつつある。

まとめ
植生の内容のとらえかたは、植物相 (Flora) 的なとらえ方と植生 (Vegetation) 帯的なとらえ方がある。植生帯的なとらえ方で問題となるのは移行帯の解釈である。中間温帯とハイマツ帯は特に研究者によっては長年議論されてきている。個人的には地域ごとに整理することで解決に近づくのではないかと思う。富士山も斜面によって違いがあるように、地域によって気候や地形は異なるからである。
また、植生帯と気候帯はセットで表現されることが多いが、世界的な規模での区分と日本の区分との違いや対応について理解が求められる。それらを踏まえて日本の重層的に形成されてきた森林植生を理解したい。
コメント